摘要:本研究從傳統(tǒng)發(fā)酵食品中篩選產(chǎn)脂肪酶乳酸菌,結(jié)合干酪漿快速成熟模型得到產(chǎn)內(nèi)酯化合物性能較好的乳酸菌,將其作為附屬發(fā)酵劑應(yīng)用于切達(dá)干酪的制備�����,采用感官評價�、氣相色譜-質(zhì)譜(ga
本研究從傳統(tǒng)發(fā)酵食品中篩選產(chǎn)脂肪酶乳酸菌��,結(jié)合干酪漿快速成熟模型得到產(chǎn)內(nèi)酯化合物性能較好的乳酸菌��,將其作為附屬發(fā)酵劑應(yīng)用于切達(dá)干酪的制備,采用感官評價���、氣相色譜-質(zhì)譜(gas chromatography-mass spectrometry, GC-MS)聯(lián)用技術(shù)比較添加產(chǎn)脂肪酶乳酸菌的附屬發(fā)酵劑和未添加附屬發(fā)酵劑干酪在4、10�、14℃成熟過程中(150 d)風(fēng)味品質(zhì)及內(nèi)酯化合物含量的變化規(guī)律�。結(jié)果表明��,產(chǎn)脂肪酶乳酸片球菌4D和格氏乳球菌Y3具有較好的產(chǎn)內(nèi)酯性能�。經(jīng)感官評價發(fā)現(xiàn)��,與未添加附屬發(fā)酵劑的切達(dá)干酪相比����,添加乳酸片球菌4D的干酪在10��、14℃成熟90 d時分別具有香氣強度最高的奶香味��、果香;添加格氏乳球菌Y3的干酪在14℃成熟120 d具有最高的奶香味香氣強度值��。利用GC-MS技術(shù)共鑒定出4種內(nèi)酯類化合物�����,添加乳酸片球菌4D和格氏乳球菌Y3的切達(dá)干酪在3種恒溫條件下成熟時,δ-癸內(nèi)酯和δ-十二內(nèi)酯的含量均高于未添加附屬發(fā)酵劑的切達(dá)干酪��,且能夠促進(jìn)δ-辛內(nèi)酯的形成����。此外,僅在格氏乳球菌Y3發(fā)酵的切達(dá)干酪中鑒定出γ-丁內(nèi)酯�����。通過探究干酪成熟溫度和時間對內(nèi)酯形成的影響��,發(fā)現(xiàn)在14℃條件下干酪的成熟效果最好���,添加乳酸片球菌4D的切達(dá)干酪在90 d時總內(nèi)酯含量達(dá)到最高�,添加格氏乳球菌Y3的切達(dá)干酪則是在120 d時達(dá)最高��。
關(guān)鍵詞:乳酸菌;脂肪酶;切達(dá)干酪;內(nèi)酯化合物;附屬發(fā)酵劑
論文《高產(chǎn)脂肪酶乳酸菌的篩選及其對切達(dá)干酪內(nèi)酯類化合物形成的影響》發(fā)表在《食品科學(xué)》����,版權(quán)歸《食品科學(xué)》所有。本文來自網(wǎng)絡(luò)平臺����,僅供參考����。
一�、引言
切達(dá)干酪是以牛奶為原料,經(jīng)濃縮��、發(fā)酵等工序制成具有獨特風(fēng)味的發(fā)酵乳制品[1]���。對于消費者而言�,切達(dá)干酪產(chǎn)品的風(fēng)味品質(zhì)是影響是否購買干酪產(chǎn)品的重要因素[2]�����。附屬發(fā)酵劑是指在干酪加工過程中除了添加主發(fā)酵劑乳酸菌外添加的一類可用于提高干酪風(fēng)味品質(zhì)或促進(jìn)成熟的選定微生物[3]�。乳酸菌因在發(fā)酵過程可產(chǎn)生乳酸促進(jìn)凝乳便于排出乳清���,還可通過碳水化合物代謝����、脂肪酸代謝形成的醛、酮、內(nèi)酯化合物賦予干酪奶香���、果香風(fēng)味而備受關(guān)注[4-5]。例如Gobbetti等[6]的研究表明,在切達(dá)干酪中附屬發(fā)酵劑可促進(jìn)蛋白水解改善其風(fēng)味品質(zhì)�����?�?祪?yōu)等[7]將乳酸片球菌(Pediococeus acidilaetici)AS185制成附屬發(fā)酵劑用于切達(dá)干酪的發(fā)酵成熟���,與僅添加商業(yè)發(fā)酵劑干酪相比����,添加附屬發(fā)酵劑的干酪具有更好感官屬性�����。
內(nèi)酯類化合物在干酪中的含量較低,但其嗅覺閾值低����,屬于干酪的關(guān)鍵風(fēng)味化合物[8]��。牛乳中的甘油三酯在脂肪酶的作用下分解成脂肪酸,并進(jìn)一步通過β-氧化及環(huán)化生成γ-或δ-內(nèi)酯類化合物[9]�����。利用微生物所產(chǎn)的脂肪酶改善產(chǎn)品風(fēng)味可滿足消費者對食品天然��、健康的需求��,因此這種方式也備受關(guān)注[10]。如研究人員發(fā)現(xiàn)將米曲霉(Aspergillus oryzae)所產(chǎn)的脂肪酶與嗜熱鏈球菌(Streptococcus thermophilus)�、鼠李糖乳酪桿菌(Lacticaseibacillus rhamnosus)等乳酸菌混合發(fā)酵�����,可顯著提高酸奶的風(fēng)味品質(zhì)[11]。洛克菲特青霉菌(Penicillium roqueforti)所產(chǎn)的脂肪酶在藍(lán)紋干酪成熟過程中能夠提高干酪的質(zhì)量��、外觀與風(fēng)味[12]�。產(chǎn)脂肪酶微生物還能夠在不影響產(chǎn)品的理化性質(zhì)情況下促進(jìn)干酪的成熟[13]。但不同微生物所產(chǎn)脂肪酶在酶學(xué)特性上存在差異[14]�����,因此探究干酪的加工參數(shù)(如溫度����、成熟時間)對于利用微生物改善干酪風(fēng)味十分重要�。目前�,國內(nèi)外研究學(xué)者發(fā)現(xiàn)瑞士乳桿菌(Lactobacillus helveticus)、干酪乳酪桿菌(Lacticaseibacillus casei)在干酪成熟14 d時可促進(jìn)其中苯甲醛、乙偶姻的形成[15]。在成熟60 d時,經(jīng)副干酪乳酪桿菌�、發(fā)酵乳桿菌發(fā)酵的干酪中乙酸�、己酸乙酯��、苯乙醇含量較高[16]。
切達(dá)干酪的成熟過程以及風(fēng)味的穩(wěn)定形成耗時較長�,因此國內(nèi)外學(xué)者通常建立干酪漿快速成熟模型以實現(xiàn)快速準(zhǔn)確評價干酪成熟過程中的風(fēng)味品質(zhì)變化[17-18]����。干酪漿體系能夠加速干酪的成熟����,如Boran等[19]利用干酪漿加速了干酪的成熟,并實現(xiàn)干酪風(fēng)味的改善��。這一加速過程是由于干酪漿具有較高的含水量和成熟溫度(30℃)�,從而加快體系內(nèi)微生物的生化代謝[20]。干酪漿快速成熟模型也可用于評估乳酸菌在干酪發(fā)酵過程的作用�����,如李春燕等[21]利用干酪漿快速成熟模型篩選出具有較好蛋白水解能力的附屬發(fā)酵劑乳酸菌���。
由于缺乏優(yōu)質(zhì)的產(chǎn)脂肪酶乳酸菌資源及其對切達(dá)干酪內(nèi)酯化合物貢獻(xiàn)尚不明晰���,因此有必要篩選高產(chǎn)脂肪酶乳酸菌�����,開發(fā)相應(yīng)的附屬產(chǎn)香發(fā)酵劑�,并探究工藝參數(shù)對干酪風(fēng)味調(diào)控的影響���?;诖耍狙芯坷萌∷岣视王ネ该魅ΨńY(jié)合銅皂法從不同發(fā)酵樣品中篩選高產(chǎn)脂肪酶乳酸菌�����,對其生長特性進(jìn)行分析��。進(jìn)一步制作干酪漿快速成熟模型探究不同高產(chǎn)脂肪酶乳酸菌的產(chǎn)內(nèi)酯類化合物性能,篩選產(chǎn)內(nèi)酯能力較高的菌株���,通過感官評價和氣相色譜-質(zhì)譜(gas chromatography-mass spectrometry, GC-MS)技術(shù)解析高產(chǎn)脂肪酶乳酸菌及其在不同成熟時間與溫度條件下對切達(dá)干酪內(nèi)酯類化合物的貢獻(xiàn),以期為開發(fā)改善切達(dá)干酪內(nèi)酯類風(fēng)味的附屬發(fā)酵劑以及促進(jìn)我國干酪產(chǎn)業(yè)發(fā)展提供理論參考��。
二��、材料與方法
2.1 材料與試劑
巴氏殺菌乳購自光明乳業(yè)股份有限公司;由乳酸乳球菌乳亞種(Lactococcus lactis subsp. lactis)和乳酸乳球菌乳脂亞種(L. lactis subsp. cremoris)組成的Choozit MA 14 LYO發(fā)酵劑購自美國國際香精香料公司���。乳餅為云南大理市售;臭豆腐鹵水為上海浦東新區(qū)市售;奶疙瘩為新疆烏魯木齊市售;奶豆腐為內(nèi)蒙古阿拉善盟市售�����。
MARZYME MT2200凝乳酶(米黑根毛霉Rhizomucor miehei酸性蛋白酶,酶活力≥2200 IMCU/g)購自美國國際香精香料公司;MRS肉湯培養(yǎng)基購自北京陸橋技術(shù)股份有限公司;正構(gòu)烷烴(C?~C??)(色譜純)購自美國Sigma-Aldrich公司;2-辛醇(分析純)購自德國Dr. Ehrenstorfer公司;吐溫-80、司盤-80(均為分析純)購自上海阿拉丁生化科技股份有限公司;DNA抽提試劑盒�、脂肪酶活性檢測試劑盒購自生工生物工程(上海)股份有限公司��。
2.2 儀器與設(shè)備
Veriti96聚合酶鏈?zhǔn)椒磻?yīng)(polymerase chain reaction, PCR)儀購自美國Applied Biosystems公司;Bio 5000Plus電泳凝膠成像分析系統(tǒng)購自上海中晶科技有限公司;TGL-16M紫外-可見分光光度計購自上海尤尼柯儀器有限公司;DC1741型干酪小型制備機購自上海承歡輕工機械有限公司;DVB/CAR/PDMS固相微萃取纖維頭購自美國Supelco公司;7890B-5977B型GC-MS儀購自美國Agilent公司。
2.3 方法
2.3.1 培養(yǎng)基的配制
- 三丁酸甘油酯培養(yǎng)基的配制:100 mL體積分?jǐn)?shù)為10%的三丁酸甘油酯、蛋白胨10 g�、牛肉粉3 g��、氯化鈉5 g、瓊脂15 g��、900 mL Tris-HCl緩沖液(pH 8.0)��,121℃滅菌20 min�。
- 產(chǎn)酶培養(yǎng)基的配制:20 g橄欖油乳化液(稱取1.4 g吐溫-80�,加入13.2 g去離子水中,邊加邊攪拌均勻����,再于80℃水浴溶解至溶液透明后取出��,加入4 g橄欖油和1.4 g司盤-80,用均質(zhì)機處理20 min至混合液呈乳白色的均勻乳狀液���,需現(xiàn)用現(xiàn)配)、蛋白胨10 g、牛肉粉10 g�、酵母粉5 g、硫酸鎂0.1 g���、醋酸鈉5 g�、檸檬酸銨2 g�����、磷酸二氫鉀2 g��、硫酸錳0.05 g、蒸餾水1000 mL,pH 7.0�����,121℃滅菌20 min���。
2.3.2 高產(chǎn)脂肪酶乳酸菌的分離篩選與鑒定
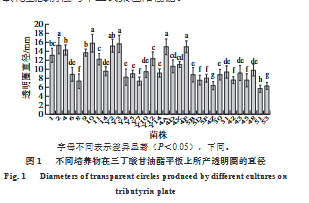
- 產(chǎn)脂肪酶乳酸菌的初篩:在無菌條件下����,從4種傳統(tǒng)發(fā)酵食品(乳餅、臭豆腐鹵水�����、奶疙瘩�、奶豆腐)中分離出外觀形態(tài)不同的單菌落,劃線純化后用30%甘油保存于-80℃。參考李曉楠等[22]的方法并稍作修改,在三丁酸甘油酯平板上打直徑為0.8 cm的小孔�,精確量取100 μL菌液接種于小孔�����,置于37℃的培養(yǎng)箱中培養(yǎng)48 h,觀察平板上是否出現(xiàn)透明圈,通過測量不同菌株所形成透明圈的直徑,初步評估菌株產(chǎn)脂肪酶能力。
- 產(chǎn)脂肪酶乳酸菌的生物學(xué)鑒定:將產(chǎn)脂肪酶菌株的單菌落接種于MRS瓊脂培養(yǎng)基,37℃培養(yǎng)24 h����,觀察并記錄菌落的大小����、形態(tài)等特征�。挑取單菌落進(jìn)行過氧化酶實驗以及革蘭氏染色并鏡檢�,記錄菌體形態(tài)。對產(chǎn)脂肪酶菌株進(jìn)行16S rDNA序列鑒定�����,采用基因組DNA抽提試劑盒提取菌體DNA��,PCR擴(kuò)增引物為27F(5’-GAGAGTTTGATCCTGGCTCAG-3’)和1492R(5’-CGGCTACCTTGTTACGACTT-3’)����。PCR擴(kuò)增條件:95℃預(yù)變性10 min;94℃變性30 s��、56℃退火30 s��、72℃延伸90 s、72℃后延伸5 min�����,30個循環(huán)��,最后4℃保存�。PCR擴(kuò)增產(chǎn)物使用1%的瓊脂糖凝膠電泳檢驗��,送往生工生物工程(上海)股份有限公司測序�。所得16S rDNA的測序結(jié)果利用NCBI數(shù)據(jù)庫中的Blast比對分析�,將與待鑒定的基因序列同源性大于99%(相似度>99%認(rèn)定為同一個種[23])的16S rDNA序列使用MEGA11軟件構(gòu)建系統(tǒng)發(fā)育樹。
- 高產(chǎn)脂肪酶乳酸菌的復(fù)篩:測定初篩得到產(chǎn)脂肪酶乳酸菌的脂肪酶活力�����,篩選高產(chǎn)脂肪酶乳酸菌����。將產(chǎn)酶乳酸菌接種于MRS培養(yǎng)基中�,置于37℃培養(yǎng)至波長600 nm處的OD值=1.0。隨后以2%的接種量分別接種到10 mL產(chǎn)酶培養(yǎng)基中��,37℃培養(yǎng)24 h��,得到對應(yīng)的發(fā)酵液�。取1 mL發(fā)酵培養(yǎng)液在7000 r/min、4℃條件下離心10 min��,收集上清粗酶液���。按照脂肪酶活性檢測試劑盒說明書測定�,按式(1)計算脂肪酶活力:
[脂肪酶活力 /(U / mL)=frac{A}{t} × F quad (1)]
式中:A為待測樣品在710 nm波長處的吸光度;t為催化反應(yīng)的時間/min;F為發(fā)酵液的稀釋倍數(shù)。
2.3.3 高產(chǎn)脂肪酶乳酸菌的生長特性評價
- 高產(chǎn)脂肪酶乳酸菌耐酸性評價:活化后的高產(chǎn)脂肪酶乳酸菌置于37℃培養(yǎng)至波長600 nm處的OD值=1.0����,以2%的接種量分別接種于pH 5.0和pH 6.0的MRS肉湯培養(yǎng)基中�,每隔3 h測定不同pH值條件下的OD?????����。
- 高產(chǎn)脂肪酶乳酸菌耐鹽性評價:活化后的高產(chǎn)脂肪酶乳酸菌置于37℃培養(yǎng)至波長600 nm處的OD值=1.0,以2%的接種量分別轉(zhuǎn)接于含不同質(zhì)量分?jǐn)?shù)(2%�����、3%和5%)NaCl的MRS肉湯培養(yǎng)基中(2%�����、3%為制備切達(dá)干酪通常的NaCl用量[24]�����,考慮到鹽分布的均勻性及極端情況下NaCl的質(zhì)量分?jǐn)?shù)會達(dá)到5%[25-26],故將該條件納入)����。于37℃恒溫培養(yǎng)箱內(nèi)培養(yǎng)24 h,每隔3 h測定菌株在不同NaCl含量MRS肉湯培養(yǎng)基中的OD?????以評價乳酸菌的耐鹽性。
- 乳酸菌的自溶度測定:以磷酸鹽緩沖液為對照�����,測定菌體懸液和對照在37℃條件下培養(yǎng)24 h的OD?????�。按式(2)計算菌體的自溶度:
[自溶度 / \%=frac{OD_{0}-OD_{24}}{OD_{0}} × 100 quad (2)]
式中:OD?為初始的OD值;OD??為培養(yǎng)24 h后的OD值。
2.3.4 附屬發(fā)酵劑的制備
取生長特性較好的高產(chǎn)脂肪酶乳酸菌接種于10 mL MRS肉湯培養(yǎng)基中���,37℃恒溫培養(yǎng)至波長600 nm處的OD值=1.0。按2%的接種量轉(zhuǎn)接于50 mL產(chǎn)酶培養(yǎng)基中�����,37℃恒溫培養(yǎng)24 h后取上述發(fā)酵液到離心管中���,在4℃����、10000 r/min條件下離心10 min,沉淀物用質(zhì)量分?jǐn)?shù)0.85%生理鹽水洗滌2次,最后將菌泥重懸于0.85%生理鹽水中得到附屬發(fā)酵劑菌液�。
2.3.5 干酪漿快速成熟模型的制作
參考郭婷等[20]的方法�。巴氏殺菌乳水浴加熱至30℃后測定pH值(pH?)����,然后向巴氏殺菌乳中加入1%(質(zhì)量分?jǐn)?shù))的商業(yè)發(fā)酵劑(乳酸乳球菌乳亞種和乳酸乳球菌乳脂亞種),并在30℃條件下發(fā)酵,在發(fā)酵過程中測定pH值(pH?)����,當(dāng)pH?與pH?間差值大于0.4時�����,向其中加入1.5%(質(zhì)量分?jǐn)?shù))的凝乳酶����,攪拌混勻后靜置40 min。等待巴氏殺菌乳凝固后,將其切割成豆腐乳樣形狀,并在切割過程中繼續(xù)水浴加熱,以0.2℃/min的速率升溫至38℃���,待析出乳清的pH值降至6.15時,排出乳清并保持溫度為38℃,當(dāng)凝乳凝結(jié)成塊后將其切割成30 cm×30 cm×30 cm的塊狀����。每隔15~20 min翻轉(zhuǎn)1次�����,重復(fù)3次�����,待排出的乳清pH值降至5.45時完成凝塊的堆積,得到干酪凝塊。隨后取100 g干酪凝塊裝入無菌真空袋中,對照組加入50 mL質(zhì)量分?jǐn)?shù)為5%的滅菌NaCl溶液����,實驗組加入47 mL質(zhì)量分?jǐn)?shù)為5%的滅菌NaCl溶液和3 mL附屬發(fā)酵劑菌液(乳酸菌濃度為10×10? CFU/mL�,添加量0.8%)攪打成漿狀�����,真空密封完成干酪漿快速成熟模型的制作�����。在30℃恒溫環(huán)境中成熟1、3、6����、9、12 d��。根據(jù)干酪漿中內(nèi)酯化合物的含量����,篩選出產(chǎn)內(nèi)酯性能較好的高產(chǎn)脂肪酶乳酸菌。
2.3.6 切達(dá)干酪的制備
參考本課題組前期建立的方法[3]�����。巴氏殺菌乳打入干酪罐中并加熱至31℃���,記錄此時的pH值�����,分別加入質(zhì)量分?jǐn)?shù)1%發(fā)酵劑(乳酸乳球菌乳亞種和乳酸乳球菌乳脂亞種)和附屬發(fā)酵劑(乳酸菌濃度為1×10¹? CFU/mL���,添加量0.8%)�����,并設(shè)置未添加附屬發(fā)酵劑的對照組,隨后在31℃條件下發(fā)酵30 min��,待殺菌乳pH值與發(fā)酵前相比下降0.4時加入質(zhì)量分?jǐn)?shù)1.5%凝乳酶���,攪拌后凝乳40 min����。隨后將凝塊切割成邊長為1 cm的方塊,切割過程中以0.1℃/min的速率熱燙至39℃����,隨后將干酪罐蓋上蓋子使溫度維持在39℃,待析出乳清的pH值為6.15時排出乳清�����,將凝乳切割成30 cm×30 cm×30 cm的方塊�����。干酪塊堆積在一起�����,每隔15 min翻轉(zhuǎn)1次,重復(fù)3次��。待析出乳清的pH值為5.45后將凝乳塊切成1 cm×1 cm×1 cm的方塊����。隨后將質(zhì)量分?jǐn)?shù)2.5%的NaCl溶液均勻撒到切達(dá)干酪上并進(jìn)行攪拌��,入模型壓榨成型并真空包裝,分別放于4����、10�、14℃(10~14℃有助于加快干酪的成熟[3,30])恒溫條件成熟30�、60、90�、120����、150 d(切達(dá)干酪的成熟期為2~36個月[30-31])��。
2.3.7 干酪的感官評價
由20名經(jīng)感官品評培訓(xùn)的評價員組成評價小組(女性10名�����、男性10名�,平均年齡24歲)。所有的感官測試按照國際標(biāo)準(zhǔn)ISO 8589:2007在感官實驗室中執(zhí)行���,室內(nèi)溫度控制在20℃。待測試的干酪樣品用數(shù)字隨機編碼�,每個干酪樣品切成大小均勻的顆粒并稱取5 g��,分別保存在一個有蓋的、無氣味的玻璃容器中(總?cè)萘繛?0 mL)。然后�,按隨機順序排列����,依次呈送給評價員進(jìn)行感官評價測試���。感官評價員按照評價表上的屬性和定義對每個干酪樣品進(jìn)行評分�����,所采用的描述詞與參考標(biāo)準(zhǔn)分別為奶香味(0.002%的丁二酮溶液)�����、果香味(0.002%的己酸乙酯溶液)、硫味(搗碎的雞蛋殼)����、酸腐味(0.1%的丁酸溶液)���、堅果味(生堅果)����、干酪味(新鮮黃油)����。在干酪香氣感官評價中�,每個香氣屬性以0~10進(jìn)行強度等級劃分(0為無強度或香氣無察覺,5為中等強度��,10為非常強)�����。每個干酪樣品重復(fù)檢驗3次�。
2.3.8 干酪的GC-MS分析
采用頂空固相微萃取法對干酪中內(nèi)酯類化合物進(jìn)行提取�����。分別精確稱量5.0 g干酪樣品���,碾碎后放入萃取瓶中���,加入100 μL 220 mg/L的2-辛醇溶液�,在60℃的恒溫水浴鍋中平衡5 min���,通過DVB/CAR/PDMS固相微萃取頭在60℃條件下萃取45 min。采用HP-INNOWAX色譜柱(60 m×0.25 mm,0.25 μm);進(jìn)樣口溫度250℃�。程序升溫:40℃保持3 min����,以5℃/min的速率升至140℃����,保持2 min,3℃/min升至240℃,保持15 min����。載氣為氦氣(純度為99.99%);流量為2 mL/min;進(jìn)樣方式為不分流進(jìn)樣。
質(zhì)譜條件:電子轟擊能量:70 eV;離子源溫度:230℃;四極桿溫度:150℃;發(fā)射電流:35 µA;掃描速率:1.9 scans/s;質(zhì)量掃描范圍m/z 30~450���。所有揮發(fā)性化合物的定性采用NIST17質(zhì)譜庫進(jìn)行檢索,選擇匹配度大于80%的物質(zhì)確定化合物��。再利用正構(gòu)烷烴C?~C??計算保留指數(shù)(retention index, RI)��,并與文獻(xiàn)報道值進(jìn)行比對��,兩者結(jié)合對干酪內(nèi)酯類化合物進(jìn)行定性分析。采用內(nèi)標(biāo)法進(jìn)行定量分析����,分別按式(3)���、(4)計算各組分含量和RI:
[內(nèi)酯類化合物含量 /(mg / kg)=frac{
ho_{0} × V_{0}}{m} × frac{S_{a}}{S_{0}} quad (3)]
式中:ρ?為內(nèi)標(biāo)物的質(zhì)量濃度/(mg/L);V?為內(nèi)標(biāo)物的體積/L;m為樣品質(zhì)量/kg;S?為待測化合物的峰面積;S?為內(nèi)標(biāo)物的峰面積����。
[RI=100 × n+100 × frac{RT-RT_{n}}{RT_{n+1}-RT_{n}} quad (4)]
式中:RT為化合物的保留時間/min;RT?和RT???分別為碳數(shù)n和n+1正構(gòu)烷烴的保留時間/min(RT?
2.4 數(shù)據(jù)處理
每個實驗重復(fù)3次��,采用Excel 2019軟件整理實驗數(shù)據(jù)�����,以±s表示。采用SPSS 26.0軟件對數(shù)據(jù)進(jìn)行顯著性分析��,利用Origin 2019b軟件繪圖�����。
三��、結(jié)果與分析
3.1 高產(chǎn)脂肪酶乳酸菌的篩選
三丁酸甘油酯平板上培養(yǎng)物周圍出現(xiàn)透明圈表明該培養(yǎng)物可產(chǎn)生降解三丁酸甘油酯(C?)中短鏈底物的脂肪酶,因此該方法常用于脂肪酶生產(chǎn)菌的篩選[32]�����。本研究采用三丁酸甘油酯透明圈法從乳餅�、臭豆腐鹵水��、奶疙瘩��、奶豆腐中篩選出33株具有透明圈形成能力的培養(yǎng)物。將初篩培養(yǎng)物經(jīng)劃線純化后在MRS瓊脂平板上的菌落形態(tài)呈白色或乳白色,菌落較小且呈圓形,邊緣光滑。經(jīng)過過氧化氫酶實驗與革蘭氏染色得到10株過氧化氫酶陰性與革蘭氏染色陽性菌���。
將菌株的16S rDNA序列上傳至NCBI數(shù)據(jù)庫進(jìn)行Blast比對,選取相似性大于99%的菌株序列,使用MEGA11軟件將其與分離菌株進(jìn)行多序列比對�����,構(gòu)建系統(tǒng)發(fā)育樹�����。結(jié)果顯示��,菌株Y3、Y2為格氏乳球菌(L. garvieae)�,4X�����、42、6為戊糖片球菌(Pediococcus pentosaceus)����,4D���、2為乳酸片球菌(P. acidilactici)�,31、43����、30為植物乳植桿菌(Lactiplantibacillus plantarum)。
隨后利用銅皂法對初篩具有產(chǎn)脂肪酶能力的乳酸菌所產(chǎn)脂肪酶的活性進(jìn)行測定,結(jié)果顯示����,從云南�、新疆����、內(nèi)蒙古等主要牧民生活地區(qū)發(fā)酵乳制品所分離的戊糖片球菌6、戊糖片球菌4X、乳酸片球菌4D、乳酸片球菌2所產(chǎn)脂肪酶具有相對較高的活力,其中乳酸片球菌4D酶活力最高,達(dá)3.32±0.54 U/mL����,格氏乳球菌Y3酶活力為1.46±0.27 U/mL�。
3.2 高產(chǎn)脂肪酶菌株的生長特性分析
3.2.1 耐酸性與耐鹽性分析
研究菌株生長環(huán)境中NaCl含量與pH值對其生長的影響情況對于切達(dá)干酪的制備具有重要意義���。在切達(dá)干酪成熟過程(60~180 d)中的pH值范圍通常在5.0~6.0之間[33]���。在pH 5.0�����、6.0的MRS培養(yǎng)基中���,6株產(chǎn)脂肪酶乳酸菌均可正常生長�����。在2%、3% NaCl溶液環(huán)境中6株產(chǎn)酶乳酸菌均能夠正常生長;當(dāng)NaCl質(zhì)量分?jǐn)?shù)達(dá)到5%時,格氏乳球菌Y2和戊糖片球菌6的生長受到抑制。這表明格氏乳球菌Y3�����、乳酸片球菌2����、戊糖片球菌4X和乳酸片球菌4D能夠較好適應(yīng)常規(guī)以及非常規(guī)切達(dá)干酪的環(huán)境���,也可用于干酪漿快速成熟模型的制作�,從而進(jìn)一步評估菌株在干酪中的應(yīng)用潛力。
3.2.2 菌株的自溶度分析
菌株的自溶度是影響干酪質(zhì)量和口味的重要因素����,它可以幫助細(xì)菌產(chǎn)生胞內(nèi)酶����,從而改善干酪的風(fēng)味[34-35]��。戊糖片球菌4X����、格氏乳球菌Y3�、乳酸片球菌4D�、乳酸片球菌2的自溶度均大于15%�。研究表明,菌株的自溶度大于15%能夠快速釋放胞內(nèi)酶進(jìn)行代謝反應(yīng)��,從而促進(jìn)風(fēng)味化合物的形成[36-37]�。綜合不同產(chǎn)脂肪酶乳酸菌的耐酸耐鹽特性,選擇戊糖片球菌4X��、格氏乳球菌Y3�����、乳酸片球菌4D、乳酸片球菌2用于建立干酪漿快速成熟模型��,分析其產(chǎn)內(nèi)酯能力�����。
3.3 基于干酪漿快速成熟模型的高產(chǎn)脂肪酶乳酸菌產(chǎn)內(nèi)酯化合物性能分析
在干酪漿的12 d成熟期內(nèi)�,由不同產(chǎn)脂肪酶乳酸菌發(fā)酵的干酪漿中內(nèi)酯總含量在成熟的第3天到第9天迅速增加��,在成熟后期又緩慢增加;乳酸片球菌4D發(fā)酵成熟的干酪漿中總內(nèi)酯含量(181.88 μg/kg)高于其他不同產(chǎn)酶乳酸菌發(fā)酵干酪漿�,且是對照組樣品(71.25 μg/kg)的2.6倍�,格氏乳球菌Y3發(fā)酵的干酪漿中總內(nèi)酯含量(157.75 μg/kg)是對照組的2.2倍。在5組干酪漿樣品中均鑒定出δ-癸內(nèi)酯和δ-十二內(nèi)酯;在乳酸片球菌4D和格氏乳球菌Y3發(fā)酵的干酪漿中還鑒定出δ-辛內(nèi)酯�,第12天的含量分別為25.78 μg/kg和39.71 μg/kg;僅在格氏乳球菌Y3發(fā)酵的干酪漿中鑒定出γ-丁內(nèi)酯(84.1 μg/kg)。本研究發(fā)現(xiàn)乳酸片球菌4D和格氏乳球菌Y3能夠豐富干酪漿快速成熟模型中的內(nèi)酯化合物種類與含量�����,具有促進(jìn)內(nèi)酯風(fēng)味形成的潛力,因此選擇這兩株乳酸菌進(jìn)一步探究產(chǎn)脂肪酶菌株對切達(dá)干酪產(chǎn)品風(fēng)味的影響作用��。
3.4 不同高產(chǎn)脂肪酶乳酸菌所制備干酪的感官評價
干酪的成熟溫度與成熟時間會改變菌株的代謝活動����,從而影響干酪的風(fēng)味品質(zhì)[38-39]����。通過對分別添加了2株產(chǎn)脂肪酶乳酸菌和未添加產(chǎn)脂肪酶乳酸菌的干酪香氣進(jìn)行感官評價�����,探究其在4�����、10℃和14℃條件下成熟30��、60、90、120��、150 d的風(fēng)味變化情況。感官評價結(jié)果表明乳酸片球菌4D和格氏乳球菌Y3發(fā)酵干酪樣品的風(fēng)味強度隨時間的延長、溫度的增加而逐漸增強���。在10、14℃成熟90 d時,添加高產(chǎn)脂肪酶乳酸菌4D和Y3干酪樣品的奶香味和果香味均高于對照組���,且在14℃條件下添加乳酸片球菌4D干酪的奶香味和果香味強度值達(dá)到最高,分別為7.9和7.4���。而含有格氏乳球菌Y3的干酪在14℃成熟120 d時��,具有最高的奶香味香氣強度,為7.9。在成熟150 d時���,不同成熟溫度下的干酪風(fēng)味強度均降低。
3.5 不同成熟時間和溫度下高產(chǎn)脂肪酶乳酸菌所制備干酪的內(nèi)酯類化合物分析
添加2種高產(chǎn)脂肪酶乳酸菌以及未添加高產(chǎn)脂肪酶乳酸菌的切達(dá)干酪在150 d成熟周期中內(nèi)酯化合物種類以及含量變化顯示,通過NIST 17.0譜庫與RI共檢出4種內(nèi)酯類化合物�����,包括δ-癸內(nèi)酯�����、δ-十二內(nèi)酯��、δ-辛內(nèi)酯和γ-丁內(nèi)酯。δ-癸內(nèi)酯是切達(dá)干酪中最為常見的內(nèi)酯化合物,賦予干酪椰子的風(fēng)味屬性[40]。對照組干酪的δ-癸內(nèi)酯含量最高達(dá)106.05 μg/kg���,添加乳酸片球菌4D和格氏乳球菌Y3的干酪最高分別可達(dá)245.33、353.24 μg/kg。δ-十二內(nèi)酯通常賦予干酪奶油以及奶香的風(fēng)味����,已有研究表明其是干酪中的關(guān)鍵風(fēng)味化合物[41]��。δ-十二內(nèi)酯在對照組成熟過程中含量最高達(dá)44.58 μg/kg,添加乳酸片球菌4D與格氏乳球菌Y3的干酪中δ-十二內(nèi)酯的最高含量分別為108.85、77.17 μg/kg���。δ-辛內(nèi)酯也是干酪中的主要內(nèi)酯�����,其對干酪的風(fēng)味有重要貢獻(xiàn)�,能夠賦予干酪椰子的風(fēng)味屬性[42-43]��。添加乳酸片球菌4D與格氏乳球菌Y3的干酪在成熟前60 d即可檢測到δ-辛內(nèi)酯�����,而對照組在成熟90 d后才能夠檢測出。此外�,僅在添加格氏乳球菌Y3的干酪樣品中檢測到γ-丁內(nèi)酯��,其可貢獻(xiàn)椰子味和奶油味的風(fēng)味屬性[44]。綜上���,產(chǎn)脂肪酶乳酸菌能夠提高干酪中內(nèi)酯化合物的含量與種類,且在所篩選的兩株高產(chǎn)脂肪酶乳酸菌中��,格氏乳球菌Y3所發(fā)酵的干酪含有較高含量的內(nèi)酯化合物與種類����,因此其具有的產(chǎn)內(nèi)酯性能更好。
影響切達(dá)干酪中內(nèi)酯化合物形成的因素包括溫度�����、水分�、附屬發(fā)酵劑以及原料乳的預(yù)處理方式[45]。本研究探究了不同成熟時間和溫度對干酪樣品中內(nèi)酯化合物形成的影響�。結(jié)果表明����,在4~14℃成熟120 d時����,切達(dá)干酪中的δ-癸內(nèi)酯含量最高����,δ-辛內(nèi)酯與δ-十二內(nèi)酯均在成熟90 d時檢測到最高值,且內(nèi)酯含量隨成熟溫度的提高而提高��。γ-丁內(nèi)酯在4~10℃成熟150 d時含量達(dá)最高(24.52~38.45 μg/kg)���,而在14℃成熟90 d時可達(dá)到最高(54.25 μg/kg)��?���?梢?����,較高的成熟溫度能夠促進(jìn)內(nèi)酯化合物的形成���。此外�����,在14℃成熟120 d后干酪中內(nèi)酯化合物含量逐漸降低或積累變慢��。因此,較高的成熟溫度雖然對干酪內(nèi)酯風(fēng)味具有積極影響���,但成熟時間不宜過長。綜上�,選擇產(chǎn)脂肪酶乳酸菌Y3發(fā)酵干酪�,且在14℃成熟120 d適于干酪內(nèi)酯化合物的形成��。
四�、討論
切達(dá)干酪中的內(nèi)酯類化合物是牛乳中甘油三酯在脂肪酶的作用下形成脂肪酸����,進(jìn)一步在β-氧化酶���、環(huán)化酶的作用下形成�����,對干酪整體風(fēng)味具有重要貢獻(xiàn)[46]����。研究發(fā)現(xiàn)�����,產(chǎn)脂肪酶乳酸乳球菌乳亞種(雙乙酰型)(L. lactis subsp. lactis biovar diacetylactis)能夠提高干酪中脂肪酸的含量[47]��。因此,將產(chǎn)脂肪酶乳酸菌作為附屬發(fā)酵劑與商業(yè)發(fā)酵劑混合可作為調(diào)控干酪內(nèi)酯類化合物生物合成的策略。干酪漿快速成熟模型因具有較高的含水量與成熟溫度�����,可用于快速評估附屬發(fā)酵劑乳酸菌的風(fēng)味形成潛力[17]���。此外�,探究成熟溫度及時間對切達(dá)干酪風(fēng)味形成的影響已成為一個熱點[3,28,48],但目前對于產(chǎn)脂肪酶乳酸菌與切達(dá)干酪內(nèi)酯化合物之間的相關(guān)性鮮有報道。本研究基于三丁酸甘油酯平板、銅皂法和干酪漿快速成熟模型篩選適用于制備切達(dá)干酪、具有較好產(chǎn)內(nèi)酯能力的高產(chǎn)脂肪酶乳酸菌并制備附屬發(fā)酵劑,結(jié)合感官評價與GC-MS探究含高產(chǎn)脂肪酶乳酸菌的附屬發(fā)酵劑以及不同成熟溫度與時間對切達(dá)干酪內(nèi)酯化合物形成的影響����。
脂肪酶能夠催化甘油三酯的水解從而形成切達(dá)干酪的風(fēng)味前體——游離脂肪酸�,通常外源添加的脂肪酶適用于原料乳��,因此利用附屬發(fā)酵劑在干酪發(fā)酵過程中產(chǎn)生脂肪酶,可避免因原料乳熱處理引起的酶失活[49]�����。乳酸菌是脂肪酶的來源之一����,如Esteban-Torres等[49]發(fā)現(xiàn)植物乳植桿菌所產(chǎn)的脂肪酶能夠降解三丁酸甘油酯。本研究從發(fā)酵乳制品所分離的戊糖片球菌6�����、戊糖片球菌4X�����、乳酸片球菌4D�、乳酸片球菌2具有相對較高的脂肪酶活力���。李曉楠等[22]結(jié)合三丁酸甘油酯平板透明圈法以及銅皂法從新疆地區(qū)發(fā)酵乳制品中篩選出高產(chǎn)脂肪酶的植物乳植桿菌��、瑞士乳酸桿菌(L. helveticus)���。此外����,從臭豆腐鹵水分離出的格氏乳球菌Y2、格氏乳球菌Y3所產(chǎn)的脂肪酶活性也相對較高。有研究表明臭豆腐鹵水中含有較多微生物所產(chǎn)生的脂肪酶[50]。目前研究的產(chǎn)脂肪酶菌株主要是從土壤�、污水等環(huán)境中篩選分離[51-52]��,本研究結(jié)果為食源產(chǎn)脂肪酶乳酸菌的篩選提供了新的參考。
干酪漿快速成熟模型與切達(dá)干酪在NaCl添加量方面存在差異,前者的NaCl添加量通常在5%����,切達(dá)干酪中的NaCl質(zhì)量分?jǐn)?shù)在2%~4%[24,27]。結(jié)合不同NaCl濃度下的生長曲線�����,發(fā)現(xiàn)格氏乳球菌Y3��、乳酸片球菌2����、戊糖片球菌4X和乳酸片球菌4D在5%的NaCl環(huán)境下能夠生長�����,因此可應(yīng)用于干酪漿快速成熟模型��。干酪漿快速成熟模型能夠縮短干酪的成熟周期[17-19]。Kristoffersen等[27]發(fā)現(xiàn)干酪漿在30℃成熟3 d后����,可形成切達(dá)干酪的獨特風(fēng)味���。將格氏乳球菌Y3��、乳酸片球菌2、戊糖片球菌4X和乳酸片球菌4D應(yīng)用于干酪漿快速成熟模型��,發(fā)現(xiàn)在干酪漿12 d的成熟期中����,內(nèi)酯化合物含量從第3天開始迅速增加。這一加速過程是由于干酪漿具有較高的含水量和成熟溫度�����,從而加快體系內(nèi)微生物的生化代謝[27]���。但有研究表明�,這種高溫�����、高水分的環(huán)境會影響體系中的化學(xué)反應(yīng)和酶催化反應(yīng)以及增加微生物腐敗等風(fēng)險���,因此干酪漿快速模型更適合篩選特定的微生物[53]�����。另一方面,乳酸菌的自溶與風(fēng)味密切相關(guān)��,如Drake等[54]通過促進(jìn)發(fā)酵劑菌株的自溶改善干酪品質(zhì)����。乳酸片球菌4D和格氏乳球菌Y3的自溶度與其他菌株相比相對較高,這可能與其較好的產(chǎn)內(nèi)酯性能相關(guān)����。
附屬發(fā)酵劑的添加對切達(dá)干酪的感官評分與風(fēng)味形成具有重要的影響[38]�。將干酪漿快速成熟模型篩選出產(chǎn)內(nèi)酯性能較好的乳酸片球菌4D和格氏乳球菌Y3制成附屬發(fā)酵劑��,并應(yīng)用于切達(dá)干酪的制備��,發(fā)現(xiàn)添加高產(chǎn)脂肪酶乳酸菌4D和Y3干酪樣品具有更濃郁的奶香味和果香味�。添加乳酸片球菌4D和格氏乳球菌Y3提高了切達(dá)干酪中δ-癸內(nèi)酯�、δ-十二內(nèi)酯的含量,且縮短了δ-辛內(nèi)酯的形成時間����,只有格氏乳球菌Y3在成熟過程中形成γ-丁內(nèi)酯��。研究表明,乳球菌能夠促進(jìn)脂肪分解并有效改善干酪風(fēng)味[55]���。李宇輝等[56]發(fā)現(xiàn)產(chǎn)脂肪酶乳酸菌附屬發(fā)酵劑能夠改善干酪的感官品質(zhì)�����,促進(jìn)具有奶油味����、果香味的風(fēng)味化合物形成�。因此產(chǎn)脂肪酶乳酸菌能夠改善切達(dá)干酪的風(fēng)味感官,促進(jìn)內(nèi)酯類化合物的形成�。
成熟溫度對切達(dá)干酪的風(fēng)味也有影響[43]��。切達(dá)干酪中的內(nèi)酯化合物含量隨成熟溫度的提高呈上升趨勢,其中14℃相比其他2種溫度能夠加快內(nèi)酯化合物的形成����,如γ-丁內(nèi)酯在4~10℃成熟150 d時含量達(dá)到最高����,而在14℃成熟90 d即可達(dá)到最高值�。這可能是由于干酪中微生物菌群的代謝酶活性被激活,從而促進(jìn)微生物代謝產(chǎn)香[57-58]。Walsh等[30]發(fā)現(xiàn)10℃成熟8個月與7℃成熟12個月的干酪感官品質(zhì)相似��,較高的成熟溫度能夠促進(jìn)干酪風(fēng)味的形成�,這與本研究結(jié)果一致。另一方面,成熟時間對內(nèi)酯的積累具有重要作用��,Alewijn等[59]發(fā)現(xiàn)γ-內(nèi)酯類化合物需要更長的成熟時間才能達(dá)到最高濃度�����,這與本研究中γ-丁內(nèi)酯的積累趨勢一致��。王姣等[48]對不同成熟周期切達(dá)干酪的揮發(fā)性化合物分析發(fā)現(xiàn),切達(dá)干酪中內(nèi)酯化合物的含量隨成熟時間的延長緩慢增加。Chen Chen等[8]對英國威克農(nóng)場切達(dá)干酪的內(nèi)酯化合物進(jìn)行分析�����,發(fā)現(xiàn)δ-辛內(nèi)酯�、δ-癸內(nèi)酯以及δ-十二內(nèi)酯在早期至成熟階段迅速積累�,隨后緩慢變化。在添加產(chǎn)脂肪酶乳酸菌附屬發(fā)酵劑的切達(dá)干酪中���,這3種內(nèi)酯化合物在成熟90~120 d達(dá)到最高值,隨后逐漸下降�����。但對于內(nèi)酯化合物與切達(dá)干酪的成熟標(biāo)志之間相關(guān)性�����,還需通過風(fēng)味指紋圖譜技術(shù)結(jié)合多元統(tǒng)計方法進(jìn)一步分析判斷�����。
五、結(jié)論
本研究利用三丁酸甘油酯透明圈法����、銅皂法并結(jié)合干酪漿快速成熟模型篩選得到耐酸耐鹽性與產(chǎn)內(nèi)酯性能較好的乳酸片球菌4D和格氏乳球菌Y3��。將乳酸片球菌4D和格氏乳球菌Y3作為附屬發(fā)酵劑用于切達(dá)干酪的制備,研究發(fā)現(xiàn)添加這兩株菌的切達(dá)干酪中內(nèi)酯化合物種類與含量均提高���,且改善了切達(dá)干酪的感官屬性。格氏乳球菌Y3能夠促進(jìn)切達(dá)干酪γ-丁內(nèi)酯的形成��,且在14℃成熟120 d的效果較好,在切達(dá)干酪的附屬發(fā)酵劑開發(fā)方面具有良好的應(yīng)用潛力。后續(xù)研究可進(jìn)一步測定干酪成熟過程中脂肪酸含量的變化�����,以探究乳酸菌所產(chǎn)脂肪酶在內(nèi)酯化合物形成過程中的作用以及內(nèi)酯化合物的形成與切達(dá)干酪成熟的相關(guān)性�����。
參考文獻(xiàn)
[1] KENDIRCI P, SALUM P, BAS D, et al. Production of enzyme-modified cheese (EMC) with ripened white cheese flavour: Ⅱ-effects of lipases[J]. Food and Bioproducts Processing, 2020, 122: 230-244. DOI:10.1016/j.fbp.2020.05.010.
[2] CHEN C, TIAN T H, YU H Y, et al. Characterisation of the key volatile compounds of commercial Gouda cheeses and their contribution to aromas according to Chinese consumers’ preferences[J]. Food Chemistry: X, 2022, 15: 100416. DOI:10.1016/j.fochx.2022.100416.
[3] 陳臣, 田同輝, 周文雅, 等. 高產(chǎn)3-甲基丁醛乳酸乳球菌對切達(dá)干酪風(fēng)味的影響[J]. 中國食品學(xué)報, 2023, 23(11): 276-288. DOI:10.16429/j.1009-7848.2023.11.027.
[4] XIA A N, YANG Y, GUO M, et al. Unveiling of the key pathway in flavor formation in fermented milk of Lactococcus lactis subsp. lactis via genomics and metabolomics[J]. Food Bioscience, 2023, 56: 103159. DOI:10.1016/j.fbio.2023.103159.
[5] JIANG N, WU R N, WU C, et al. Multi-omics approaches to elucidate the role of interactions between microbial communities in cheese flavor and quality[J]. Food Reviews International, 2023, 39(8): 5446-5458. DOI:10.1080/87559129.2022.2070199.
[6] GOBBETTI M, DE ANGELIS M, DI CAGNO R, et al. Pros and cons for using non-starter lactic acid bacteria (NSLAB) as secondary/adjunct starters for cheese ripening[J]. Trends in Food Science & Technology, 2015, 45(2): 167-178. DOI:10.1016/j.tifs.2015.07.016.
[7] 康優(yōu), 呂楊, 高磊, 等. 乳酸片球菌與馬克斯克魯維酵母混合附屬發(fā)酵劑對切達(dá)干酪品質(zhì)的影響[J/OL]. 食品科學(xué), 2024 (2024-09-19). https://kns.cnki.net/kcms/detail/11.2206.TS.20240918.0959.039.html.
[8] CHEN C, LIU Z, YU H Y, et al. Flavoromic determination of lactones in cheddar cheese by GC-MS-olfactometry, aroma extract dilution analysis, aroma recombination and omission analysis[J]. Food Chemistry, 2022, 368: 130736. DOI:10.1016/j.foodchem.2021.130736.
[9] SUTAY KOCABA? D, LYNE J, USTUNOL Z. Hydrolytic enzymes in the dairy industry: applications, market and future perspectives[J]. Trends in Food Science & Technology, 2022, 119: 467-475. DOI:10.1016/j.tifs.2021.12.013.
[10] BORA L, GOHAIN D, DAS R. Recent advances in production and biotechnological applications of thermostable and alkaline bacterial lipases[J]. Journal of Chemical Technology & Biotechnology, 2013, 88(11): 1959-1970. DOI:10.1002/jctb.4170.
[11] HUANG Y Y, YU J J, ZHOU Q Y, et al. Preparation of yogurt-flavored bases by mixed lactic acid bacteria with the addition of lipase[J]. LWT-Food Science and Technology, 2020, 131: 109577. DOI:10.1016/j.lwt.2020.109577.
[12] CARON T, LE PIVER M, PÉRON A C, et al. Strong effect of Penicillium roqueforti populations on volatile and metabolic compounds responsible for aromas, flavor and texture in blue cheeses[J]. International Journal of Food Microbiology, 2021, 354: 109174. DOI:10.1016/j.ijfoodmicro.2021.109174.
[13] RANI S, JAGTAP S. Acceleration of Swiss cheese ripening by microbial lipase without affecting its quality characteristics[J]. Journal of Food Science and Technology, 2019, 56(1): 497-506. DOI:10.1007/s13197-018-3482-6.
[14] 楊萬齡, 沈興旺, 崔鳳怡, 等. 藍(lán)紋牦牛干酪成熟特性及其風(fēng)味分析[J]. 食品科學(xué)技術(shù)學(xué)報, 2023, 41(6): 52-64. DOI:10.12301/spxb202300008.
[15] XIANG Q, XIA Y X, CHEN L, et al. Flavor precursors and flavor compounds in Cheddar-flavored enzyme-modified cheese due to pre-enzymolysis combined with lactic acid bacteria fermentation[J]. Food Bioscience, 2023, 53: 102698. DOI:10.1016/j.fbio.2023.102698.
[16] RANDAZZO C L, LIOTTA L, ANGELIS M, et al. Adjunct culture of non-starter lactic acid bacteria for the production of provola Dei nebrodi PDO cheese: in vitro screening and pilot-scale cheesemaking[J]. Microorganisms, 2021, 9(1): 179. DOI:10.3390/microorganisms9010179.
[17] CASTADA H Z, HARPER W J, BARRINGER S A. Volatile organic compounds of a Swiss cheese slurry system with and without added reduced glutathione, compared with commercial Swiss cheese[J]. International Dairy Journal, 2015, 49: 72-77. DOI:10.1016/j.idairyj.2015.04.010.
[18] DARNAY L, MIKLÓS G, L?RINCZ A, et al. Possible inhibitory effect of microbial transglutaminase on the formation of biogenic amines during Trappist cheese ripening[J]. Food Additives & Contaminants. Part A, Chemistry, Analysis, Control, Exposure & Risk Assessment, 2022, 39(3): 580-587. DOI:10.1080/19440049.2021.2005831.
[19] BORAN O S, SULEJMANI E, HAYALOGLU A A. Acceleration of proteolysis, flavour development and enhanced bioactivity in a model cheese using Kuflu cheese slurry: an optimisation study[J]. Food Chemistry, 2023, 412: 135495. DOI:10.1016/j.foodchem.2023.135495.
[20] 郭婷, 余志堅, 陳超, 等. 基于快速成熟模型的藏靈菇發(fā)酵切達(dá)干酪揮發(fā)性風(fēng)味物質(zhì)分析[J]. 食品科學(xué), 2018, 39(8): 90-96. DOI:10.7506/spkx1002-6630-201808015.
[21] 李春燕, 熊智強, 王光強, 等. 乳酸菌附屬發(fā)酵劑的篩選及其對干酪漿蛋白水解的影響[J]. 食品與發(fā)酵工業(yè), 2022, 48(17): 129-135. DOI:10.13995/j.cnki.11-1802/ts.030574.
[22] 李曉楠, 李宇輝, 盧士玲, 等. 新疆傳統(tǒng)奶酪中產(chǎn)脂肪酶乳酸菌的優(yōu)選及酶學(xué)特性研究[J]. 食品與發(fā)酵工業(yè), 2021, 47(10): 50-57. DOI:10.13995/j.cnki.11-1802/ts.025978.
[23] 李斯淼, 陳芳敏, 方麗, 等. 一株家蠶腸道分離乳酸菌的鑒定及生物學(xué)特性[J/OL]. 微生物學(xué)通報. https://doi.org/10.13344/j.microbiol.china.240512.
[24] 吳怡霏, 劉宗尚, 劉維娜, 等. NaCl對原制干酪品質(zhì)影響及減鈉方法研究進(jìn)展[J]. 中國食品添加劑, 2022, 33(5): 186-193. DOI:10.19804/j.issn1006-2513.2022.05.025.
[25] ONG L, D’INCECCO P, PELLEGRINO L, et al. The effect of salt on the structure of individual fat globules and the microstructure of dry salted Cheddar cheese[J]. Food Biophysics, 2020, 15(1): 85-96. DOI:10.1007/s11483-019-09606-x.
[26] MISTRY V V, KASPERSON K M. Influence of salt on the quality of reduced fat Cheddar cheese 1[J]. Journal of Dairy Science, 1998, 81(5): 1214-1221. DOI:10.3168/jds.S0022-0302(98)75681-4.
[27] KRISTOFFERSEN T, MIKOLAJCIK E M, GOULD I A. Cheddar cheese flavor. IV. directed and accelerated ripening process[J]. Journal of Dairy Science, 1967, 50(3): 292-297. DOI:10.3168/jds.s00220302(67)87412-5.
[28] 莫蓓紅, 趙建, 苗君蒞, 等. 一種植物乳桿菌對干酪模型成熟中微生物及蛋白質(zhì)水解的影響[J]. 食品工業(yè)科技, 2012, 33(7): 100-104. DOI:10.13386/j.issn1002-0306.2012.07.062.
[29] MUEHLENKAMP-ULATE M R, WARTHESEN J J. Evaluation of several nonstarter lactobacilli for their influence on Cheddar cheese slurry proteolysis 1[J]. Journal of Dairy Science, 1999, 82(7): 1370-1378. DOI:10.3168/jds.S0022-0302(99)75362-2.
[30] WALSH E A, DIAKO C, SMITH D M, et al. Influence of storage time and elevated ripening temperature on the chemical and sensory properties of white Cheddar cheese[J]. Journal of Food Science, 2020, 85(2): 268-278. DOI:10.1111/1750-3841.14998.
[31] 程晶晶, 陳會民, 羅潔, 等. 不同成熟期切達(dá)干酪的組分及特征風(fēng)味物質(zhì)的解析[J]. 食品與發(fā)酵工業(yè), 2020, 46(15): 231-237. DOI:10.13995/j.cnki.11-1802/ts.023149.
[32] GARCÍA-CANO I, ROCHA-MENDOZA D, KOSMERL E, et al. Purification and characterization of a phospholipid-hydrolyzing phosphoesterase produced by Pediococcus acidilactici isolated from Gouda cheese[J]. Journal of Dairy Science, 2020, 103(5): 3912-3923. DOI:10.3168/jds.2019-17965.
[33] XIA Y J, YUAN R, WENG S N, et al. Proteolysis, lipolysis, texture and sensory properties of cheese ripened by Monascus fumeus[J]. Food Research International, 2020, 137: 109657. DOI:10.1016/j.foodres.2020.109657.
[34] ZHAO S Q, DORAU R, TØMMERHOLT L, et al. Simple & better-accelerated cheese ripening using a mesophilic starter based on a single strain with superior autolytic properties[J]. International Journal of Food Microbiology, 2023, 407: 110398. DOI:10.1016/j.ijfoodmicro.2023.110398.
[35] KANG O J, VÉZINZ L P, LABERGE S, et al. Some factors influencing the autolysis of Lactobacillus bulgaricus and Lactobacillus casei[J]. Journal of Dairy Science, 1998, 81(3): 639-646. DOI:10.3168/jds.s0022-0302(98)75618-8.
[36] AYAD E H E, NASHAT S, EL-SADEK N, et al. Selection of wild lactic acid bacteria isolated from traditional Egyptian dairy products according to production and technological criteria[J]. Food Microbiology, 2004, 21(6): 715-725. DOI:10.1016/j.fm.2004.02.009.
[37] TONDHOUSH A, SOLTANI M, AZARIKIA F, et al. Fabrication of UF-white cheese: Obtaining a different proteolysis rate, texture, and flavor via using combinations of mesophilic starter culture and Lactobacillus helveticus[J]. Food Science & Nutrition, 2023, 12(1): 328-339. DOI:10.1002/fsn3.3769.
[38] BASSI D, GAZZOLA S, SATTIN E, et al. Lactic acid bacteria adjunct cultures exert a mitigation effect against spoilage microbiota in fresh cheese[J]. Microorganisms, 2020, 8(8): 1199. DOI:10.3390/microorganisms8081199.
[39] SIHUFE G A, ZORRILLA S E, PEROTTI M C, et al. Acceleration of cheese ripening at elevated temperature. an estimation of the optimal ripening time of a traditional Argentinean hard cheese[J]. Food Chemistry, 2010, 119(1): 101-107. DOI:10.1016/j.foodchem.2009.06.001.
[40] DELGADO F J, GONZÁLEZ-CRESPO J, CAVA R, et al. Formation of the aroma of a raw goat milk cheese during maturation analysed by SPME-GC-MS[J]. Food Chemistry, 2011, 129(3): 1156-1163. DOI:10.1016/j.foodchem.2011.05.096.
[41] HIGH R, BREMER P, KEBEDE B, et al. Comparison of four extraction techniques for the evaluation of volatile compounds in spray-dried New Zealand sheep milk[J]. Molecules, 2019, 24(10): 1917. DOI:10.3390/molecules24101917.
[42] PLUTA-KUBICA A, DOMAGA?A J, G?SIOR R, et al. Characterisation of the profile of volatiles of Polish Emmental cheese[J]. International Dairy Journal, 2021, 116: 104954. DOI:10.1016/j.idairyj.2020.104954.
[43] QIAN M, REINECCIUS G. Identification of aroma compounds in Parmigiano-Reggiano cheese by gas chromatography/olfactometry[J]. Journal of Dairy Science, 2002, 85(6): 1362-1369. DOI:10.3168/jds.S0022-0302(02)74202-1.
[44] FAN S S, TANG K, XU Y, et al. Characterization of the potent odorants in Tibetan Qingke Jiu by sensory analysis, aroma extract dilution analysis, quantitative analysis and odor activity values[J]. Food Research International, 2020, 137: 109349. DOI:10.1016/j.foodres.2020.109349.
[45] 于海燕, 姚文倩, 陳臣, 等. 內(nèi)酯類化合物在食品中的風(fēng)味貢獻(xiàn)及形成機制[J]. 現(xiàn)代食品科技, 2022, 38(5): 337-349; 55. DOI:10.13982/j.mfst.1673-9078.2022.5.0954.
[46] 陳臣, 劉政, 于海燕, 等. 奶酪中內(nèi)酯類物質(zhì)風(fēng)味貢獻(xiàn)及其生物合成調(diào)控進(jìn)展[J]. 現(xiàn)代食品科技, 2020, 36(11): 305-312. DOI:10.13982/j.mfst.1673-9078.2020.11.0513.
[47] TUNGJAROENCHAI W, WHITE C H, HOLMES W E, et al. Influence of adjunct cultures on volatile free fatty acids in reduced-fat Edam cheeses[J]. Journal of Dairy Science, 2004, 87(10): 3224-3234. DOI:10.3168/jds.S0022-0302(04)73458-X.
[48] 王姣, 許凌云, 張晉華, 等. 不同成熟時間切達(dá)奶酪中揮發(fā)性香氣成分及其電子鼻判別分析[J]. 食品科學(xué), 2020, 41(20): 175-183. DOI:10.7506/spkx1002-6630-20190726-349.
[49] ESTEBAN-TORRES M, MANCHEÑO J M, DE LAS RIVAS B, et al. Production and characterization of a tributyrin esterase from Lactobacillus plantarum suitable for cheese lipolysis[J]. Journal of Dairy Science, 2014, 97(11): 6737-6744. DOI:10.3168/jds.2014-8234.
[50] TIAN H X, ZOU L, LI L, et al. Characterisation of the aroma profile and dynamic changes in the flavour of stinky tofu during storage[J]. Foods, 2023, 12(7): 1410. DOI:10.3390/foods12071410.
[51] 許琳琳, 劉慧乾, 張夢瑤, 等. 低溫脂肪酶產(chǎn)生菌的篩選、表達(dá)及酶學(xué)性質(zhì)分析[J]. 食品工業(yè)科技, 2024, 45(20): 133-140. DOI:10.13386/j.issn1002-0306.2023110093.
[52] 王娜, 趙文倩, 張盈盈, 等. 產(chǎn)脂肪酶多粘類芽胞桿菌的產(chǎn)酶條件和固定化優(yōu)化及對模擬油污廢水的降解研究[J]. 微生物學(xué)雜志, 2024, 44(4): 31-39. DOI:10.3969/j.issn.1005-7021.2024.04.004.
[53] FOX P F, WALLACE J M, MORGAN S, et al. Acceleration of cheese ripening[J]. Antonie van Leeuwenhoek, 1996, 70(2/3/4): 271-297. DOI:10.1007/BF00395937.
[54] DRAKE M A, BOYLSTON T D, SPENCE K D, et al. Chemical and sensory effects of a Lactobacillus adjunct in Cheddar cheese[J]. Food Research International, 1996, 29(3/4): 381-387. DOI:10.1016/0963-9969(96)00006-3.
[55] KLEIN N, LORTAL S. Attenuated starters: an efficient means to influence cheese ripening: a review[J]. International Dairy Journal, 1999, 9(11): 751-762. DOI:10.1016/S0958-6946(99)00155-7.
[56] 李宇輝, 王俊鋼, 劉成江, 等. 產(chǎn)脂肪酶乳酸菌對新疆傳統(tǒng)奶酪脂肪酸及風(fēng)味的影響[J]. 農(nóng)業(yè)工程學(xué)報, 2022, 38(6): 319-329.
[57] UEDA S, SASAKI R, NAKABAYASHI R, et al. Exploring the lipids involved in the formation of characteristic lactones in Japanese black cattle[J]. Metabolites, 2021, 11(4): 203. DOI:10.3390/metabo11040203.
[58] SAHINGIL D, HAYALOGLU A A, SIMSEK O, et al. Changes in volatile composition, proteolysis and textural and sensory properties of white-brined cheese: effects of ripening temperature and adjunct culture[J]. Dairy Science & Technology, 2014, 94(6): 603-623. DOI:10.1007/s13594-014-0185-2.
[59] ALEWIJN M, SLIWINSKI E L, WOUTERS J T M. Production of fat-derived (flavour) compounds during the ripening of Gouda cheese[J]. International Dairy Journal, 2005, 15(6/7/8/9): 733-740. DOI:10.1016/j.idairyj.2004.09.009.




