摘要:為探究2,4-表油菜素內(nèi)酯(EBL)對谷子萌發(fā)的影響及其生理機制,從而為提高谷子耐鹽性提供理論支持,研究10^-10、10^-9、10^-8、10^-7 mol/L 2,4-表油菜素內(nèi)酯浸種處理對0、50、100、150、200、250、300 m
為探究2,4-表油菜素內(nèi)酯(EBL)對谷子萌發(fā)的影響及其生理機制,從而為提高谷子耐鹽性提供理論支持,研究10^-10、10^-9、10^-8、10^-7 mol/L 2,4-表油菜素內(nèi)酯浸種處理對0、50、100、150、200、250、300 mmol/L NaCl脅迫下的谷子萌發(fā)和生長的影響。結果表明,隨著NaCl脅迫濃度的升高,每組EBL浸種處理均降低了谷子發(fā)芽勢、發(fā)芽率、胚根長、胚軸長及萌發(fā)后的含水量。在0、50、100mmol/L的低濃度NaCl脅迫下,每組EBL浸種處理對谷子萌發(fā)情況均無顯著影響;在150、200mmol/L的中等濃度NaCl脅迫下,不同EBL浸種處理會不同程度地影響谷子萌發(fā)情況,在所有處理組中,10^-9 mol/L EBL對200mmol/L NaCl脅迫下谷子的萌發(fā)效果影響最顯著(P<0.05);在250、300 mmol/L的高濃度NaCl脅迫下,每組EBL浸種處理促進谷子萌發(fā)的效果均不明顯。在200 mmol/L NaCl處理下10^-9 mol/L EBL浸種處理組中,萌發(fā)谷子體內(nèi)的丙二醛(MDA)含量相比不施用EBL的對照組顯著降低(P<0.05),超氧化物歧化酶(SOD)、過氧化物酶(POD)活性相比不施用EBL的對照組顯著升高(P<0.05)。
關鍵詞: 谷子; 油菜素內(nèi)酯; 鹽脅迫; 種子萌發(fā); 生理機制
論文《油菜素內(nèi)酯浸種對鹽脅迫谷子萌發(fā)的影響及其生理機制》發(fā)表在《江蘇農(nóng)業(yè)科學》,版權歸《江蘇農(nóng)業(yè)科學》所有。本文來自網(wǎng)絡平臺,僅供參考。
正文
引言
土地鹽漬化能使得作物生長發(fā)育受到嚴重影響,是全球關注的環(huán)境問題之一,已成為干旱、半干旱地區(qū)農(nóng)業(yè)發(fā)展的主要威脅。目前,全世界鹽漬化土壤面積已經(jīng)超過10億hm²,預計到2050年,全球有50%的耕地發(fā)生鹽漬化[1]。我國鹽漬化土壤分布廣泛,總面積位居世界第3,屬于我國主要的中低產(chǎn)土壤類型之一,在很大程度上限制了我國農(nóng)業(yè)的生產(chǎn)與發(fā)展。谷子(Setaria italica)是狗尾草屬一年生草本植物,為五谷雜糧之首,谷子經(jīng)去殼處理后,成為日常生活中必不可少的小米。小米不僅有保健身體的作用,還有幫助食用者安神的作用,食用價值較高,因此,人們對于小米的需求量較大[2]。
另外,谷子除了具有較高的營養(yǎng)價值外,還具有糧飼兼用、抗旱耐瘠薄、適應性強等特點[3],是一種典型的環(huán)境友好型作物[4]。雖然谷子的耐鹽性高于水稻、玉米等作物,但是隨著我國土壤鹽漬化問題日益嚴重,對我國谷子的萌發(fā)、生長都造成較大的不利影響[5]。面對日益擴大的鹽堿地面積和巨大的糧食需求,提高鹽漬地上谷子的萌發(fā)率顯得尤為重要。
油菜素內(nèi)酯(brassinolide, BR)被列為第六大植物激素,在植物的花粉、種子及根、莖、葉中都有分布[6],BR中蕓苔素內(nèi)酯(BL)、2,4-表油菜素內(nèi)酯(EBL)和2,8-高油菜素內(nèi)酯(HBL)是活性較高的3種,油菜素內(nèi)酯作為一種新型、高效的植物激素,在植物的生長發(fā)育過程以及植物響應生物、非生物脅迫過程中發(fā)揮著不可或缺的作用,它能夠通過促進植物光合作用和抗逆性來促進植物的生長發(fā)育,影響果實產(chǎn)量和品質(zhì)形成[7]。
權夢萍等研究油菜素內(nèi)酯對非生物逆境脅迫下植物生長的影響,發(fā)現(xiàn)油菜素內(nèi)酯可以通過改善植物的相關生理系統(tǒng)來提高植物對非生物脅迫的耐受性,促進植物生長[8]。有研究者分析了油菜素內(nèi)酯對鹽脅迫下黑麥草、菜豆、綠豆和水稻種子萌發(fā)及生理的影響,發(fā)現(xiàn)外源BR處理可以提高黑麥草種子的發(fā)芽率、發(fā)芽勢,可以增強相應酶活性[9-12]。呂宗環(huán)等使用不同濃度的NaCl溶液對10份谷子進行處理,明確了適合谷子萌發(fā)期抗鹽鑒定的NaCl濃度為180 mmol/L[13]。張永芳等設置了5個碳酸鈉濃度對4個谷子品種進行鹽脅迫處理,結果表明,低濃度的碳酸鈉能夠促進谷子萌發(fā),但是隨著其濃度增加,萌發(fā)指標會逐漸降低[14]。Nie等研究2,4-表油菜素內(nèi)酯對碳酸氫鈉脅迫黃瓜種子萌發(fā)的改善作用,發(fā)現(xiàn)EBL處理后顯著提高了超氧化物歧化酶(SOD)、過氧化氫酶(CAT)、過氧化物酶(POD)的活性,緩解鹽脅迫引起的氧化損傷,顯著抵消對種子萌發(fā)的抑制作用[15]。Chakma等用2,4-表油菜素內(nèi)酯(EBL)單獨處理或?qū)⑵渑c其他激素聯(lián)合處理棉花種子進行萌發(fā)試驗,結果表明,EBL在有或無鹽脅迫的條件下都能促進種子萌發(fā),而其他激素在脅迫下均無效[16]。有研究者分析油菜素內(nèi)酯浸種對臭椿種子、豇豆種子萌發(fā)的影響發(fā)現(xiàn),浸種后其種子發(fā)芽率、發(fā)芽勢均提高,發(fā)芽速度縮短,根長和芽長、根干重和芽干重均增加,種子產(chǎn)量提高[17-18]。
目前,已有報道發(fā)現(xiàn)油菜素內(nèi)酯在其他植物中有效促進了種子萌發(fā),但尚未見油菜素內(nèi)酯對谷子萌發(fā)影響機制方面的研究,因此研究油菜素內(nèi)酯浸種對鹽脅迫下谷子萌發(fā)的影響及其生理機制具有重大意義。本研究通過油菜素內(nèi)酯浸種對鹽脅迫谷子萌發(fā)的影響試驗,以期找到最適的EBL浸種施用方式,為谷子在鹽漬地上的高產(chǎn)種植提供理論依據(jù)和實踐參考,從而促進我國農(nóng)業(yè)的可持續(xù)發(fā)展。
1 材料與方法
1.1 試驗地點與材料
試驗于2024年2月在呂梁學院植物生理實驗室進行,供試材料為晉谷21號,購買自山西鑫騰達農(nóng)業(yè)科技有限公司。
1.2 試驗方法
1.2.1 谷子挑選及浸種預處理
將無水乙醇稀釋為75%的乙醇溶液,供谷子消毒使用。配制油菜素內(nèi)酯母液,并稀釋為10^-7、10^-8、10^-9、10^-10 mol/L濃度,供谷子浸種使用。稱取NaCl,配制成50、100、150、200、250、300 mmol/L的NaCl溶液,供谷子鹽脅迫使用。對晉谷21號的種子進行粗挑選,去掉干癟的籽粒,將留下的谷子在75%乙醇中浸泡20s,用蒸餾水洗滌3次,再用吸水紙吸干谷子表面水分,分別使用配制好的不同濃度的油菜素內(nèi)酯溶液浸種處理24h,依次表示為EBL1、EBL2、EBL3、EBL4,以蒸餾水浸種24h為對照組,表示為EBL0。
1.2.2 谷子萌發(fā)培養(yǎng)
在浸種處理24h后,將浸泡谷子的浸種液倒掉,將谷子置于濾紙上,在室內(nèi)自然晾干。在直徑為9cm的玻璃培養(yǎng)皿中放置雙層濾紙,在處理好的培養(yǎng)皿中分別加入不同濃度的NaCl溶液,濃度分別為0(CK,蒸餾水)、50、100、150、200、250、300 mmol/L,每個試驗設置3次重復。進一步挑選籽粒飽滿、大小一致的谷子各100粒分別置于上述培養(yǎng)皿中,保持NaCl溶液始終浸沒種子高度的一半。最后,將上述處理好的種子放置于人工氣候箱中,于28℃黑暗條件下培養(yǎng),培養(yǎng)期間始終保持濾紙濕潤。谷子萌發(fā)后3d測定其發(fā)芽勢,發(fā)芽后7d測定其發(fā)芽率,同時進行相關指標的計算。
1.3 指標的測定
1.3.1 發(fā)芽勢、發(fā)芽率的測定
以根、芽長度均超過谷子直徑的一半作為發(fā)芽標準,從發(fā)芽后2d開始每天記錄種子發(fā)芽數(shù),將發(fā)芽后3d的發(fā)芽數(shù)用于計算發(fā)芽勢。基于發(fā)芽后7d的發(fā)芽數(shù)計算發(fā)芽率。發(fā)芽勢=發(fā)芽后3d的發(fā)芽數(shù)/供試谷子粒數(shù)×100%;發(fā)芽率=發(fā)芽后7d的發(fā)芽數(shù)/供試谷子粒數(shù)×100%。
1.3.2 胚根、胚軸長度的測定
谷子發(fā)芽7d后,從每個培養(yǎng)皿中隨機選取5粒萌發(fā)的谷子,用剛尺分別測量其胚根、胚軸長度,每個處理重復3次。結果取平均值。
1.3.3 萌發(fā)后含水量的測定
谷子萌發(fā)后7d,取每個處理濃度的谷子10粒,用吸水紙吸干種子表面殘留水分,稱取谷子鮮重(FW),在電熱鼓風干燥箱中于105℃烘干至恒重,稱得干重(DW)。萌發(fā)谷子含水量=[(FW-DW)/FW]×100%。每個處理重復3次,結果取平均值。
1.3.4 丙二醛(MDA)含量、抗氧化酶活性的測定
丙二醛含量用硫代巴比妥酸法測定;超氧化物歧化酶活性用氮藍四唑還原法測定;過氧化物酶活性用愈創(chuàng)木酚比色法測定[19]。
試驗過程中測得的各數(shù)據(jù)均用 Excel 2021、SPSS27.0進行處理、統(tǒng)計分析等操作,數(shù)據(jù)均表示為“平均數(shù)±標準差”格式,采用Duncan's新復極差測驗法(α=0.05)進行單因素顯著性方差分析。
2 結果與分析
2.1 不同處理對谷子萌發(fā)的調(diào)控效應
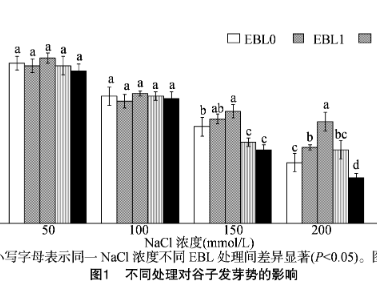
由圖1可知,隨著NaCl脅迫濃度的升高,每組EBL浸種處理均降低了谷子的發(fā)芽勢。在無NaCl脅迫的條件下,EBL3浸種處理(即10^-8 mol/L EBL處理)的谷子發(fā)芽勢較其他濃度EBL處理組高,但差異不顯著。在50、100mmol/L NaCl脅迫下,EBL2浸種處理(即10^-9 mol/L EBL處理)的谷子發(fā)芽勢比其他濃度EBL處理組高,但差異均不顯著。在150 mmol/L NaCl脅迫下,與EBL0浸種處理相比,隨EBL浸種濃度的上升,谷子發(fā)芽勢首先表現(xiàn)為升高趨勢,其中EBL2浸種處理(即10^-9 mol/L EBL處理)的發(fā)芽勢達最高;隨著EBL浸種濃度進一步上升,谷子發(fā)芽勢降低;EBL0~EBL2浸種處理與EBL3、EBL4處理組相比,對谷子發(fā)芽勢的促進效果達到顯著水平(P<0.05)。在200mmol/L NaCl脅迫下,與EBL0浸種處理相比,EBL1~EBL3浸種處理(即10^-10~10^-8 mol/L EBL處理)谷子的發(fā)芽勢均升高,EBL4浸種處理(即10^-7 mol/L EBL處理)的谷子發(fā)芽勢降低;在EBL2浸種處理(即10^-9 mol/L EBL處理)下,谷子發(fā)芽勢達到最大值,且與其他EBL處理組間均有顯著差異(P<0.05)。在250 mmol/L NaCl脅迫下,與EBL0浸種處理相比,隨著EBL浸種濃度的上升,谷子的發(fā)芽勢首先表現(xiàn)為升高趨勢,其中EBL2浸種處理(即10^-9 mol/L EBL處理)的谷子發(fā)芽勢達到最大值;隨著EBL浸種濃度進一步升高,谷子的發(fā)芽勢降低;在EBL0~EBL2浸種處理(即0、10^-10、10^-9 mol/L EBL處理)下,與EBL3、EBL4處理組相比,對谷子的發(fā)芽勢均起到了促進效果。300 mmol/L NaCl脅迫下,各組EBL浸種處理均對谷子的發(fā)芽勢無顯著影響。
由圖2可知,隨著NaCl脅迫濃度的升高,各EBL浸種處理均降低了谷子的發(fā)芽率。在無NaCl脅迫處理下,EBL1浸種處理(即10^-10 mol/L EBL處理)的谷子發(fā)芽率高于其他EBL處理,但差異不顯著。在50 mmol/L NaCl脅迫下,EBL2浸種處理(即10^-9 mol/L EBL處理)的谷子發(fā)芽率高于其他濃度EBL處理,但差異不顯著。在100 mmol/L NaCl溶液處理下,各EBL浸種處理均對谷子發(fā)芽率無顯著影響。在150 mmol/L NaCl脅迫下,EBL1~EBL3浸種處理(即10^-10~10^-8 mol/L EBL處理)的谷子發(fā)芽率高于其他處理;在EBL2浸種處理(即10^-9 mol/L EBL處理)下,谷子的發(fā)芽率達到最大值;在EBL4浸種處理(即10^-7 mol/L EBL處理)下,谷子發(fā)芽率降低;EBL1~EBL3處理組與EBL4處理組相比,對谷子發(fā)芽率的促進效果均達到顯著水平(P<0.05)。在200mmol/L NaCl脅迫下,與EBL0浸種處理相比,EBL1~EBL3浸種處理(即10^-10~10^-8 mol/L EBL處理)均可提高谷子的發(fā)芽率;在EBL4浸種處理(即10^-7 mol/L EBL處理)下,谷子發(fā)芽率較其他處理降低;在EBL2浸種處理(即10^-9 mol/L EBL處理)下,谷子的發(fā)芽率達到最大值,且此處理與其他處理相比差異顯著(P<0.05),谷子發(fā)芽率分別比EBL0、EBL1、EBL3、EBL4處理組提高32.76%、17.24%、12.07%和50.00%。在250 mmol/L NaCl脅迫下,與EBL0浸種處理相比,隨EBL浸種濃度的上升,谷子發(fā)芽率升高,EBL1、EBL2浸種處理(即10^-10、10^-9 mol/L EBL處理)谷子發(fā)芽率較其他處理差異顯著(P<0.05),但這2組處理間差異不顯著;隨著EBL浸種濃度繼續(xù)上升,谷子發(fā)芽率降低,EBL3浸種處理(即10^-8 mol/L EBL處理)對谷子發(fā)芽率的抑制作用與EBL0處理相比差異不顯著,EBL4浸種處理(即10^-7 mol/L EBL處理)較EBL0處理顯著抑制了谷子發(fā)芽率(P<0.05)。在300mmol/L NaCl脅迫下,各EBL浸種處理均對谷子發(fā)芽率無顯著影響。
2.2 不同處理對谷子胚根、胚軸長度的調(diào)控效應
由圖3可知,隨著NaCl脅迫濃度的升高,各EBL浸種處理整體上降低了谷子萌發(fā)期間的胚根長度。在無NaCl脅迫處理下,各EBL浸種處理對谷子萌發(fā)期間的胚根長度無顯著影響。在50 mmol/L NaCl脅迫處理下,EBL2浸種處理(即10^-9 mol/L EBL處理)谷子萌發(fā)期間的胚根長度較其他EBL處理組高(除EBL0浸種處理外),但差異不顯著。在100mmol/L NaCl脅迫下,EBL2浸種處理(即10^-9 mol/L EBL處理)谷子萌發(fā)期間的胚根長度大于其他EBL處理組,但差異不顯著。在150 mmol/L NaCl脅迫下,與EBL0浸種處理相比,EBL1~EBL3浸種處理(即10^-10、10^-9、10^-8 mol/L EBL處理)均增加了谷子萌發(fā)期間的胚根長度;在EBL2浸種處理(即10^-9 mol/L EBL處理)下,谷子胚根長度達到最大值;與其他處理相比,EBL4浸種處理(即10^-7 mol/L EBL處理)降低了谷子萌發(fā)期間的胚根長度。在200mmol/L NaCl脅迫下,與EBL0浸種處理相比,EBL1~EBL3浸種處理(即10^-10、10^-9、10^-8 mol/L EBL處理)均增加了谷子萌發(fā)期間的胚根長度;EBL4浸種處理(即10^-7 mol/L EBL處理)降低了谷子萌發(fā)期間的胚根長度;在EBL2浸種處理(即10^-9 mol/L EBL處理)下,谷子的胚根長度達到最大值,且此處理組與除EBL3處理組以外的其他處理組相比均有顯著差異(P<0.05)。在250mmol/L NaCl脅迫下,EBL2浸種處理(即10^-9 mol/L EBL處理)谷子萌發(fā)期間的胚根長度高于其他EBL處理組,與EBL4浸種處理(即10^-7 mol/L EBL處理)間差異顯著(P<0.05)。在300 mmol/L NaCl脅迫下,EBL1、EBL2浸種處理(即10^-10、10^-9 mol/L EBL處理)谷子萌發(fā)期間的胚根長度大于其他EBL處理,與EBL4浸種處理(即10^-7 mol/L EBL處理)差異顯著(P<0.05)。
由圖4可知,隨著NaCl脅迫濃度的升高,各EBL浸種處理整體上均降低了谷子萌發(fā)期間的胚軸長度。在無NaCl脅迫處理下,EBL3浸種處理(即10^-8 mol/L EBL處理)谷子萌發(fā)期間的胚軸長度大于其他EBL處理,但差異不顯著。在50 mmol/L NaCl脅迫下,EBL2浸種處理(即10^-9 mol/L EBL處理)谷子萌發(fā)期間的胚軸長度大于其他EBL處理,但差異不顯著。在100 mmol/L NaCl脅迫下,EBL2、EBL3浸種處理(即10^-9、10^-8 mol/L EBL處理)谷子萌發(fā)期間的胚軸長度大于其他EBL處理,但差異不顯著。在150、200 mmol/L NaCl脅迫處理下,與EBL0浸種處理相比,EBL1~EBL3浸種處理(即10^-10、10^-9、10^-8 mol/L EBL處理)谷子萌發(fā)期間的胚軸長度均增加,EBL4浸種處理(即10^-7 mol/L EBL處理)谷子萌發(fā)期間的胚軸長度均降低;在EBL2浸種處理(即10^-9 mol/L EBL處理)下,谷子胚軸長度均達到最大值,且各EBL浸種處理對谷子萌發(fā)期間的胚軸長度有不同程度的影響。在250mmol/L NaCl脅迫處理下,各EBL浸種處理對谷子萌發(fā)期間的胚軸長度無顯著影響。在300 mmol/L NaCl脅迫下,EBL1浸種處理(即10^-10 mol/L EBL處理)谷子萌發(fā)期間的胚軸長度較其他EBL處理組顯著增加(P<0.05)。
由圖3、圖4可知,在同一NaCl濃度脅迫下,隨著EBL浸種濃度的上升,谷子萌發(fā)期間胚根和胚軸長度均先上升后下降,且胚根長度的降低幅度比胚軸長度更明顯,即胚根比胚軸對鹽脅迫下EBL處理更敏感。
2.3 不同處理組對谷子萌發(fā)后含水量的調(diào)控效應
由圖5可知,隨著NaCl脅迫濃度的升高,各EBL浸種處理整體上均降低了谷子萌發(fā)后的含水量。在0、50 mmol/L NaCl脅迫處理下,與EBL0浸種處理谷子萌發(fā)后的含水量最低,但不同處理間的差異不顯著;在EBL3浸種處理(10^-8 mol/L EBL處理)下,萌發(fā)后含水量達到最大值。在100 mmol/L NaCl脅迫下,與EBL0浸種處理相比,EBL2浸種處理(即10^-9 mol/L EBL處理)谷子萌發(fā)后的含水量提高,但變化不顯著。在150 mmol/L NaCl脅迫處理下,與EBL0浸種處理相比,EBL1~EBL3處理(即10^-10、10^-9、10^-8 mol/L EBL浸種處理)均增加了谷子萌發(fā)后的含水量,EBL4浸種處理(即10^-7 mol/L EBL處理)降低了谷子萌發(fā)后的含水量,其中在EBL2浸種處理(即10^-9 mol/L EBL處理)下,谷子萌發(fā)后的含水量達到最大值,并且此處理組與除EBL3外的其他處理組相比差異顯著(P<0.05)。在200 mmol/L NaCl脅迫下,與EBL0浸種處理相比,EBL1~EBL3浸種處理(10^-10~10^-8 mol/L EBL處理)谷子萌發(fā)后的含水量均增加,EBL4浸種處理(即10^-7 mol/L EBL處理)谷子萌發(fā)后的含水量較其他處理降低,其中EBL2浸種處理(即10^-9 mol/L EBL處理)下的含水量達到最大值,且此處理與其他處理間的差異均顯著(P<0.05)。在250、300mmol/L NaCl脅迫下,EBL2浸種處理(即10^-9 mol/L EBL處理)谷子萌發(fā)后含水量整體上高于其他濃度EBL處理,但差異不顯著。
對以上試驗測得的谷子發(fā)芽勢、發(fā)芽率、萌發(fā)期間胚根長度、胚軸長度、萌發(fā)后的含水量等5個指標進行綜合分析,結果顯示,EBL2浸種處理(即10^-9 mol/L EBL處理)對200mmol/L NaCl脅迫下谷子萌發(fā)的影響最顯著。
2.4 最佳處理對谷子丙二醛含量、抗氧化酶活性的影響
由圖6可知,NaCl脅迫處理下谷子萌發(fā)后的MDA含量(以谷子萌發(fā)后的鮮重為基礎計算,下同)表現(xiàn)出顯著增加的趨勢;與NaCl脅迫處理相比,在NaCl脅迫條件下用EBL2浸種的谷子MDA含量降低,差異達到顯著水平(P<0.05)。
由圖7、圖8可知,與對照組相比,EBL2浸種處理的萌發(fā)谷子的POD活性顯著上升(P<0.05),而SOD活性差異不顯著。在NaCl脅迫下,萌發(fā)谷子的2種酶活性均顯著上升;與NaCl脅迫相比,在NaCl脅迫條件下施用EBL2浸種處理的谷子SOD、POD活性均呈現(xiàn)顯著上升的趨勢(P<0.05)。
3 結論與討論
本試驗過程中,不同濃度的NaCl脅迫均不同程度地抑制谷子的萌發(fā)效果,通過在NaCl脅迫前進行不同濃度的EBL浸種處理,對谷子萌發(fā)的影響呈現(xiàn)不同變化。在0、50、100mmol/L等低濃度NaCl脅迫下,施用不同濃度的EBL對谷子的發(fā)芽勢、發(fā)芽率、萌發(fā)期間胚根長度、胚軸長度、萌發(fā)后的含水量5個指標整體有一定的增加,但增加趨勢均不顯著;在150、200 mmol/L等中等濃度NaCl脅迫下,施用不同濃度的EBL對谷子萌發(fā)情況呈現(xiàn)不同程度的影響。綜合分析可知,10^-9 mol/L EBL浸種處理對200 mmol/L NaCl脅迫下谷子萌發(fā)的促進效果最好;250 mmol/L高濃度NaCl脅迫下,施用不同濃度的EBL對谷子發(fā)芽勢、發(fā)芽率的影響有一定的差異,但對谷子萌發(fā)期間的胚軸長度以及萌發(fā)后的含水量均無顯著差異;在300mmol/L極高濃度NaCl脅迫下,施用不同濃度的EBL對谷子萌發(fā)情況影響均不大。對試驗所得的促進谷子萌發(fā)效果最好的NaCl濃度、EBL浸種濃度單獨作用于谷子,測定其丙二醛含量、超氧化物歧化酶和過氧化物酶活性,得出NaCl脅迫下,MDA含量較高,超氧化物歧化酶、過氧化物酶活性較低,而用EBL浸種處理后,MDA含量會降低,相應的抗氧化物酶活性會升高。
范翠枝等做了不同濃度的油菜素內(nèi)酯浸種對不同濃度的NaCl脅迫番茄種子萌發(fā)的試驗,測定了番茄種子萌發(fā)期相應抗氧化代謝指標,結果表明,10^-9 mol/L EBL浸種對150 mmol/L NaCl脅迫下的番茄種子萌發(fā)效果最好,促進了SOD、POD活性的上升[20]。本試驗結果與上述研究結果[20]一致,本試驗結果得出的最適NaCl脅迫濃度與上述研究結果不一致,是由于不同物種對鹽的耐受性不同,谷子是耐鹽作物,耐鹽濃度比番茄高。趙旭慶等研究了不同NaCl脅迫、不同油菜素內(nèi)酯浸種對鹽溶液處理下黑果枸杞種子萌發(fā)的影響,發(fā)現(xiàn)低濃度的NaCl能促進種子的萌發(fā),高濃度處理則會抑制種子萌發(fā);適宜濃度EBL浸種能促進種子萌發(fā),但濃度過高時可能會有抑制作用;0.05mg/L EBL對促進其種子萌發(fā)效果最明顯[21]。本試驗結果與上述研究結果存在不一致的地方,本試驗得出,低濃度的NaCl對谷子萌發(fā)沒有表現(xiàn)出顯著的促進或抑制效應,可能是由于不同物種對低濃度NaCl脅迫的反應不同或配制的NaCl溶液濃度不同導致結論不同,具體的原因值得后續(xù)進一步研究。
參考文獻:
[1] Yue J M, Fu Z Y, Zhang L, et al. The positive effect of different 24-epiBL pretreatments on salinity tolerance in Robinia pseudoacacia L. seedlings[J]. Forests, 2019, 10(1): 4.
[2] 吳雙. 谷子栽培及病蟲害防治技術[J]. 現(xiàn)代農(nóng)村科技, 2024(4): 49-50.
[3] 張笛, 苗興芬, 王雨婷. 100份谷子品種資源萌發(fā)期耐鹽性評價及耐鹽品種篩選[J]. 作物雜志, 2019(6): 43-49.
[4] Diao X M. Production and genetic improvement of minor cereals in China[J]. The Crop Journal, 2017, 5(2): 103-114.
[5] Zeng P, Zhu P W, Qian L F, et al. Identification and fine mapping of qGR6.2, a novel locus controlling rice seed germination under salt stress[J]. BMC Plant Biology, 2021, 21(1): 36.
[6] 陳晨, 程大偉, 李蘭, 等. 油菜素內(nèi)酯調(diào)控植物耐鹽機理研究進展[J]. 中國農(nóng)業(yè)科技導報, 2024, 26(2): 1-12.
[7] 鄭婷, 程建徽, 魏靈珠, 等. 油菜素內(nèi)酯及其在園藝植物中的研究進展[J/OL]. 分子植物育種, 2024: 1-9[2024-04-15]. http://kns.cnki.net/kcms/detail/46.1068.s.20220113.1105.004.html.
[8] 權夢萍, 徐佳慧, 尹佳茗, 等. 油菜素內(nèi)酯調(diào)控植物響應非生物逆境脅迫的生理機制[J]. 植物保護學報, 2023, 50(1): 22-31.
[9] 王丹, 劉亞西, 周揚, 等. 油菜素內(nèi)酯對鹽脅迫下黑麥草種子萌發(fā)及幼苗生長的生理調(diào)控作用[J]. 草業(yè)科學, 2021, 38(6): 1110-1118.
[10] 侯會云. 油菜素內(nèi)酯對鹽脅迫下水稻種子萌發(fā)的影響[J]. 熱帶農(nóng)業(yè)科學, 2020, 40(7): 1-6.
[11] Semida W M, Rady M M. Pre-soaking in 24-epibrassinolide or salicylic acid improves seed germination, seedling growth, and anti-oxidant capacity in Phaseolus vulgaris L. grown under NaCl stress[J]. The Journal of Horticultural Science and Biotechnology, 2014, 89(3): 338-344.
[12] Rathinavelu S. Impact of plant growth regulators and nutrient consortium on growth, superoxide dismutase, Na+/K+ ratio and yield of blackgram under salinity stress[J]. International Journal of Agriculture, Environment and Biotechnology, 2018, 11(5): 791-798.
[13] 呂宗環(huán), 韓康妮, 杜曉芬, 等. 谷子萌發(fā)期耐鹽種質(zhì)鑒定及應用[J]. 植物科學學報, 2024, 42(1): 75-84.
[14] 張永芳, 宋喜娥, 王潤梅, 等. Na2CO3脅迫對谷子種子萌發(fā)的影響[J]. 種子, 2015, 34(11): 94-97.
[15] Nie W J, Gong B, Geng B, et al. The effects of exogenous 2,4-epibrassinolide on the germination of cucumber seeds under NaHCO3 stress[J]. Plants, 2024, 13(3): 394.
[16] Chakma S P, Chileshe S M, Thomas R, et al. Cotton seed priming with brassinosteroid promotes germination and seedling growth[J]. Agronomy, 2021, 11(3): 566.
[17] Li K R, Zhang W B, Li H K. Effect of natural brassinolide on germination of Ailanthus altissima seeds[J]. Forestry Studies in China, 2005, 7(2): 12-14.
[18] Fariduddin Q, Hayat S, Ali B, et al. Effect of 2,8-homobrassinolide on the growth and yield of vigna[J]. Annals of Agri-Bio Research, 2005, 21(1): 1-6.
[19] 高俊鳳. 植物生理學實驗指導[M]. 北京: 高等教育出版社, 2006: 210-217.
[20] 范翠枝, 吳馨怡, 關欣, 等. 油菜素內(nèi)酯浸種對鹽脅迫番茄種子萌發(fā)的影響及其生理機制[J]. 生態(tài)學報, 2021, 41(5): 1857-1867.
[21] 趙旭慶, 徐珊珊, 李強, 等. 鹽脅迫下24-表油菜素內(nèi)酯浸種對黑果枸杞種子萌發(fā)的影響[J]. 林業(yè)科技通訊, 2021(3): 59-61.




